
栽培大豆(Glycine max (L.) Merr.)于5000年前由野生大豆(G. soja Sieb. & Zucc.)驯化而来,是世界重要的粮油兼用作物,种子富含植物油和蛋白质。
近日,皇冠APP下载官网徐敏副教授、付爱根教授课题组联合安徽省农业科学院黄志平研究员课题组在The Plant Journal上合作发表了研究论文“Structural variation of mitochondrial genomes sheds light on evolutionary history of soybeans”, 通过研究大豆种群mtDNA的结构多样性,探讨了大豆种群mtDNA的主要构型、驯化过程中mtDNA的演化历史以及mtDNA遗传多样性和品种改良相关的可能性。

栽培大豆品种“Williams 82”(Wm82)的核基因组是大豆中第一个发布的参考基因组,也是大豆基因功能研究最常用的参考基因组,但其完整的胞质基因组尚未报道。因而,该研究通过高通量测序和Southern Blot/PCR技术,组装了Wm82的mtDNA(Wm82_mtDNA),填补了其基因组中的一个空缺。结果显示,Wm82_mtDNA由两个独立的环型染色体(370,871-bp Chr-m1与62,661-bp Chr-m2)组成。
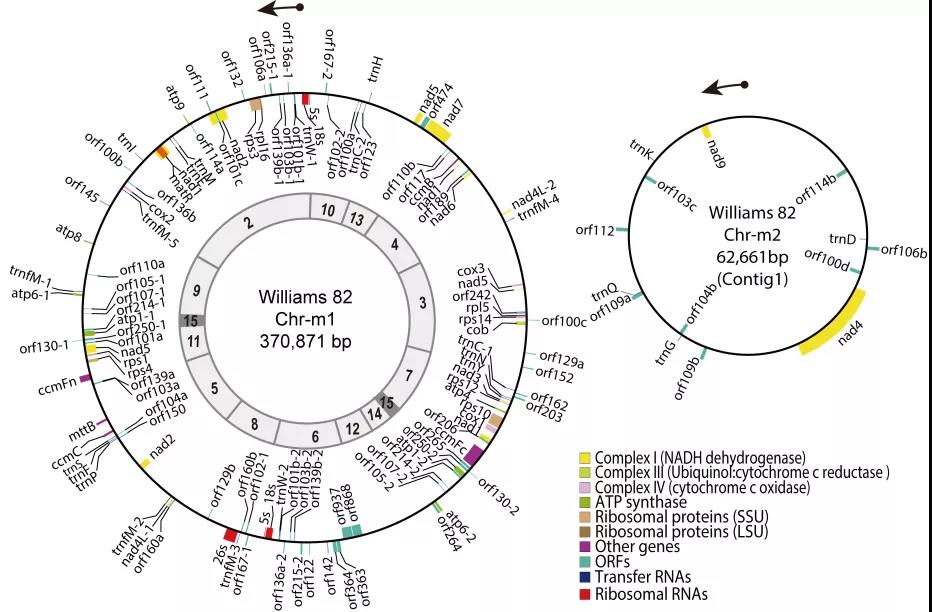
线粒体基因组(大环内圈显示的是组装中的contigs)
Wm82_mtDNA与曾报道的大豆“矮杆黄”单环mtDNA(AGH_mtDNA)有很大的结构差异,具体表现为序列的重组和得失,但是两个基因组的核心基因组成与序列差别不大。进一步分析表明,短重复片段(SR)介导的低频异位重组是单、双环构型相互转换的关键,也是大豆mtDNA结构演化的重要遗传基础。
接着,该研究利用结构变异(SV)分子标记,通过PCR/Southern blot实验或同等效力的序列分析研究了39份G. soja和182份G. max的mtDNA遗传多样性。结果,所分析的两个群体均具有5种胞质类群,分别为GSa-GSe和G1-G5,而Wm82和矮杆黄分属于G1和G5(图2)。其中,栽培大豆G1-G4类群在1992年(Grabau 等)曾报道过。
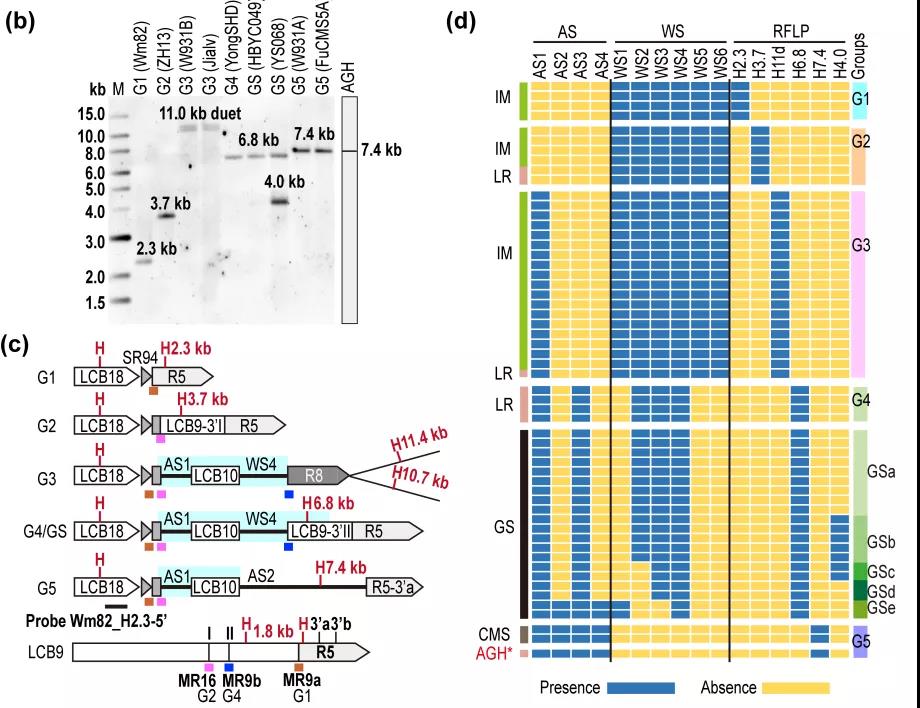
基于mtDNA结构变异对大豆胞质进行分类
研究还显示,栽培大豆中的主要胞质类型为G1-G3,占比90%以上(图2,3),与1992年报道的结果相似;而野生大豆中的胞质主要类型为GSa和GSb。它们之中只有GSe和G5胞质群的mtDNA是单环构型,其他的mtDNA均为双环型、具有保守的Chr-m2染色体,因此双环结构是大豆线粒体基因组的主要构型(图3)。
最后,通过对大豆mtDNA结构变异和叶绿体核心基因的序列差异进行系统发育分析,研究人员发现栽培大豆G1-G3的成员聚类在一个分枝上,与野生大豆胞质亲缘关系很远,其中G1 mtDNA由G3 mtDNA演化而来;而G4和G5的成员则分别与野生大豆的GSa和GSe的成员聚在一个分枝上。这充分说明了,栽培大豆主要胞质类型来源于一个祖先,与栽培大豆来源于单一驯化事件相符;而G4和G5类胞质则是通过种间杂交分别从GSa和类似GSe的野生大豆获得。因此,现代的栽培豆的胞质至少有三个不同来源。
更为有趣的是,研究还发现G3类胞质和由其演化产生的G1类胞质在改良品种(IM)中的占比(86%)相较于其在地方品种或早期栽培品种(LR)中的占比(58%)明显升高(图3b),说明胞质的遗传结构与大豆改良密切相关,但这个相关性有多少为mtDNA贡献,还需进一步研究。
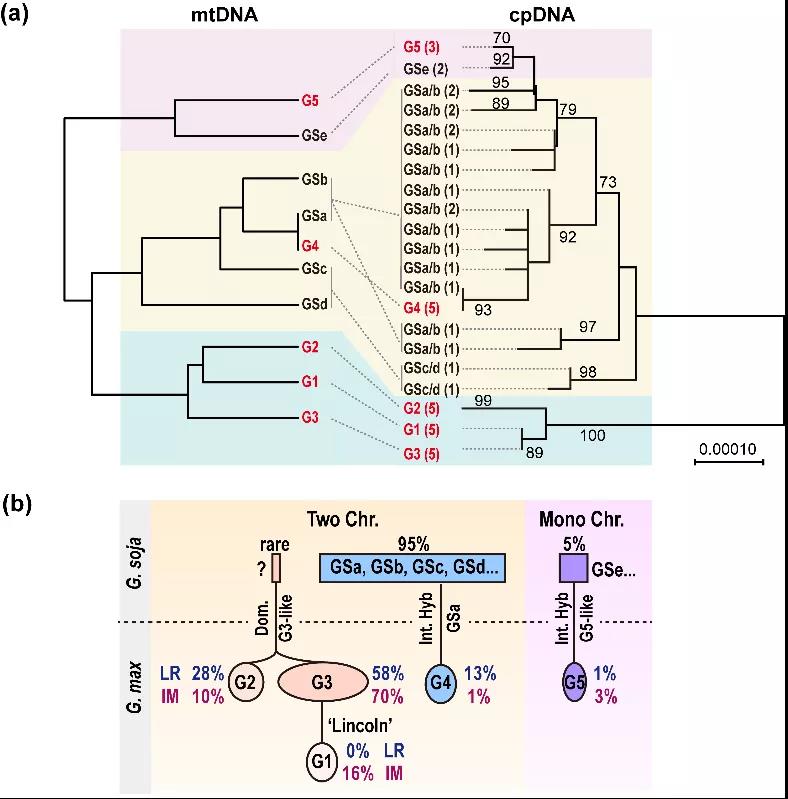
大豆胞质的聚类分析和演化历史
综上所述,该研究为大豆mtDNA遗传多样性分析及大豆基因功能分析提供了高质量的参考序列,为栽培大豆mtDNA的演化历史提供了新的见解,为胞质遗传结构影响大豆改良提供了证据。皇冠APP下载官网博士研究生刘昊为论文第一作者,徐敏副教授,付爱根教授和安徽省农业科学院黄志平研究员为该论文共同通讯作者。安徽省农业科学院王大刚副研究员和安徽省阜阳市农业科学院李智研究员参与了该研究。本研究得到国家重点研发项目,陕西省教育厅重点实验室研究项目和陕西省科技项目的资助。